


Sample collection and whole genome sequencing
58 canids from around the world were gathered for this study. This collection includes 12 gray wolves from across the Eurasian continent, 11 indigenous dogs from southern East Asia, 12 indigenous dogs from northern East Asia, 4 village dogs from Africa (Nigeria) and a set of 19 diverse dog breeds distributed across the Old World and the Americas.
Chinese indigenous dogs are dogs living in the countryside of China16 (Supplementary information, Data S1 and Figure S1) and were sampled across the geographic range of rural China, including many remote regions in Yunnan and Guizhou in southern China (Supplementary information, Table S1). The breeds include dogs from Central Asia (Afghan Hound) and North Africa (Sloughi), Europe (eight different breeds), the Arctic and Siberia (Greenland dog, Alaska Malamute, Samoyed, Siberian Husky, and East Siberian Laika), the New World (Chihuahua, Mexican and Peruvian naked dog) as well as the Tibetan Plateau (Tibetan Mastiff). These dogs were chosen to cover as many major geographic regions as possible (Figure 1A and Supplementary information, Table S1).
Population structure and genetic diversity of 58 canids. (A) Geographic locations of the 58 canids sequenced in this study. (B) Amount of of SNPs and small indels called in this study. (C) Genetic diversity for the 58 canids. AF, African village dogs; BEM, Belgian Malinois; CHI, Chihuahua; FIL, Finnish Lapphund; GAL, Galgo; GNE, Gray Norwegian Elkhound; GSD, German Shepherd Dog; JAM, Jamthund; LAH, Lapponian Herder; MEN, Mexican naked (hairless); PEN, Peruvian naked (hairless); SWL, Swedish Lapphund; AFG, Afghan Hound; SLO, Sloughi; SAM, Samoyed; ESL, East Siberian Laika; SIH, Siberian Husky; ALM, Alaska Malamute; GRD, Greenland dogs; TIM, Tibetian Mastiff. (D) Structure analysis of the 58 canids. (E) Genetic diversity of the different groups. AF, African village dogs; EB, European breeds; SI, southern Chinese indigenous dogs; W, wolves. (F) Linkage disequilibrium patterns for the different groups. (G) Principle component analysis of the 58 canids. Inset is for all individuals and the large panel is for dogs only. (H) Principle component plot for a large collection of canids together with our data. (I) A clock-like tree (UPGMA) for all the 58 individuals56.
After DNA extraction, individual genomes were sequenced to an average of 15× coverage (Supplementary information, Table S1). Of the 58 individuals, 4 gray wolves and 6 dogs have been sequenced in a previous study10. DNA sequence analysis was done using the Genome Analysis Toolkit17. After stringent filtering, we identified 20 353 184 SNPs and 3 856 246 small indels (Figure 1B), most of which are shared between groups. For example, 40.3% of the SNPs are shared between wolves, indigenous dogs and dog breeds, reflecting their recent divergence (Figure 1B). Using Sanger sequencing, we verified that the sequencing strategy was highly sensitive (false negative rate around 10%) and the amount of false positives was less than 5% (Supplementary information, Data S2 and Figure S2).
Genetic diversity and population structure
Comparison of the two haploid genomes within each individual yields the genetic diversity θ (4 Nμ) for the 58 individuals. As shown in Figure 1C, genetic diversity shows a decreasing trend from wolves to Chinese indigenous dogs (preserving 78% of the wolf heterozygosity) and subsequently to dog breeds (66% of the wolf heterozygosity), with the African village dogs having a genetic diversity comparable to many dog breeds (69% of the wolf heterozygosity). Among the dog breeds, the levels of variation in genetic diversity are quite dramatic. For example, the East Asian breed Tibetan Mastiff and East Siberian Laika show levels of diversity comparable to the Chinese indigenous dogs, but many of the European dog breeds have considerably reduced genetic diversity. Such dramatic differences in genetic diversity can be influenced both by ancient and recent history of inbreeding.
To explore the genetic relationships among these individuals, we performed a structure analysis using an expectation maximization (EM) algorithm to cluster the individuals into different numbers of groupings18. When partitioning the individuals into two groups, the algorithm separates the dogs from the wolves, with very limited admixture observed (Figure 1D). Further dividing the individuals into three subsets split the dogs into two clusters, with indigenous dogs from southern East Asia representing one subset and the other subset consisting of dog breeds from Europe and South/Central America and the African village dogs. Indigenous dogs from northern China and dog breeds from the Arctic and Central Asia, the Middle East and North Africa show a mixture of these components with varying proportions. This observation implies that there are two divergent groups of dogs: one is East Asian component and the other, non-East Asian component. It is important to emphasize that individuals with mixed constituents identified in the structure analysis are not always due to true admixture events, since populations of intermediate genotypes between these two groups tend to display mixed components (e.g., originated shortly after the split of two clades, Supplementary information, Data S3 and Figure S3). Further partitioning into four and five groups leads to the separation of the African village dogs and the breed dogs from the eastern Arctic regions (i.e., Siberian Husky, Alaska Malamute and the Greenland dog).
Genetic diversity among individuals (Figure 1C) may be heavily influenced by ancient as well as recent history, e.g., breeding programs during the last few thousand years or the past few hundred years. However, combined information from multiple breeds may reveal information about the ancestral populations that gave rise to them, since each breed has experienced separate breeding history. We therefore calculated the genetic diversity (θπ) for the “pure groups” informed by the structure analysis (K = 4, Figure 1D). As shown in Figure 1E, dog breeds, most of which of European origin, carry lower diversity than the Chinese indigenous dogs as a group, but have higher genetic diversity than the African indigenous dogs. This suggests that the ancestral population that gave rise to the European breeds was larger than the ancestral population of the African indigenous dogs. Linkage disequilibrium patterns also show similar trends (Figure 1F).
Principle component and phylogenetic analysis
When projecting the genotypes into a two-dimensional space using a principle component analysis (PCA)19, all dogs cluster together tightly compared with the distribution seen for wolves (Figure 1G, inset). When inspecting the distribution among dogs, we find that dogs spread along three major geographic axes: southern East Asia, Europe and Africa. The northern Chinese indigenous dogs and dog breeds from the Middle East/Arctic regions/Tibet fall between these three extremes (Figure 1G). The observed pattern reflects the overall geographic locations of these groups following a clear East-West gradient, which matches quite well the observation from our structure analysis.
Combining our dataset with data from a previous SNP array study, which included a larger number of samples20, we found that the southern Chinese indigenous dogs together with several East Asian dogs (e.g., Chow Chow, Akita, Chinese Shar-Pei) are closest to wolves (Figure 1H). When the phylogenetic relationships among our 58 samples are inspected, East Asian dogs spread over both sides of the deepest node connecting all dogs, while dogs from other continental areas coalesce into a subclade and then join with East Asian dogs. Thus, East Asian dogs are the most basal lineages connecting to gray wolves (Figure 1I). It is worth pointing out that the genomes of dogs from Oceania (dingoes and New Guinea singing dogs), although being closer to wolves in the PCA plot (Figure 1H), bear strong signals of admixture with gray wolves6, which likely reflects their past history of admixture, before they migrated to Australia and New Guinea (Supplementary information, Data S4 and Figure S4).
Admixture analysis
Using the joint allele frequencies among all populations in our study, we infer the split and admixture history among groups of populations using TreeMix21. If migration tracks are not allowed, then the relationships inferred from the TreeMix analysis (Figure 2A) directly reflect the patterns observed in our previous analyses including the structure (Figure 1D), the phylogenetic (Figure 1I) and the principal component analyses (Figure 1G). Thus, following the divergence between contemporary wolves and domestic dogs, the first partition within dogs is between the southern Chinese indigenous group and all other dogs. This is then followed by branching of the other dogs, largely matching the geographical distance from southern East Asia: first, dogs from Central Asia, northern China, and eastern Arctic, followed by dogs in Africa, the Middle East, and western Arctic, and the final group including all dog breeds in Europe and South/Central America.
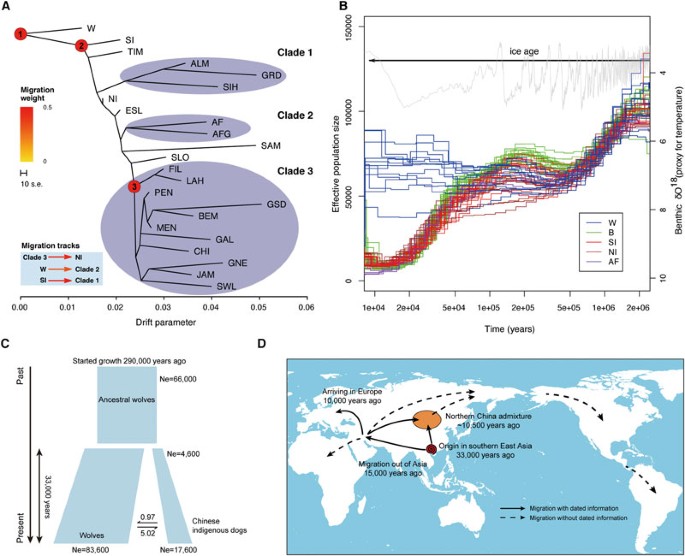
Demographic and migration histories for the domestic dog. (A) Tree topology inferred from TreeMix when no migratory tracts are allowed. The drift parameter is the amount of genetic drift along each population. Further inferred migratory tracts are shown in the bottom-left corner of the panel. The three important nodes are those that we have provided extensive dating information. (B) The PSMC plot for all the individuals. Gray lines plot the benthic δO18 levels, which are a proxy for global temperature61. The span of the current ice age (Quaternary ice age, 2.58M-now) is shown with an arrow. The x-axis is time plotted in log scale and the y-axis is effective population size. (C) Inferred population demographic history between wolves and southern East Asian indigenous dogs using the joint site frequency spectra. (D) A proposed migratory history for domestic dogs across the world based on the evidence from our study. Solid arrows represent migratory tracts that we have dating information, while dashed arrows indicate those without accurate dating.
If migration tracks are allowed in TreeMix, there is strong statistical support for migrations among a few groups: (1) northern Chinese indigenous dogs show strong admixture from European dogs (Figure 2A and Supplementary information, Data S5, Figure S5, Tables S2 and S3); (2) gene flow from wolves to the African/Middle Eastern dogs (Supplementary information, Figure S5); (3) migratory tracks from the southern Chinese dogs to the eastern Arctic group (i.e., Siberian Husky, Alaska Malamute and the Greenland dog; Supplementary information, Figure S5). When all possible migration events in the history of these samples are examined using the F3/F4 test22, there is again a strong statistical support for all the migration events listed above (Supplementary information, Data S5).
Long-term evolutionary trajectories for wolves and dogs
Using the divergence between the two haploid genomes within individuals, the pairwise sequentially Markovian coalescent (PSMC) model provides a method for investigating the long-term trajectories in population sizes23. To translate demographic history into real-time units, estimation of an accurate mutation rate is very important. Previously, several different mutation rates were used, but they were generally not carefully calibrated (Supplementary information, Data S6)24. Using multiple outgroup species to the dog (e.g., horse and cat), our estimate of the mutation rate for the lineage leading to the domestic dog is 2.2 × 10−9 per site per year (Supplementary information, Data S6 and Table S4), a rate similar to those from several earlier studies25,26. Using this mutation rate, we estimate dates for the population history of dogs and wolves. As shown in Figure 2B, a decrease in the size of the ancestral wolf population started to occur 2 million years ago, reaching a saddle point about 3-400 000 years ago. The ancestral population then increased in size, peaking at around 200 000 years ago. After a subsequent small decline in population size, wolves and dogs started to diverge from each other between 20 000 and 100 000 years ago (see next section for a more precise dating). Although all domestic dogs drastically decreased in population size after the population split, the wolf population experienced a slight growth, possibly as a consequence of the megafauna extinctions (i.e., late Quaternary extinction)27 that provided gray wolves with better food resources due to reduced competition from other predators.
Time of divergence between contemporary wolves and dogs
Treemix and phylogenetic analyses identified southern Chinese indigenous dogs as the most basal population compared to wolves, from which all other dog populations diverged. We therefore used joint allele frequencies between the 12 gray wolves and the 11 southern Chinese indigenous dogs, to infer the demographic history for these two populations with the dadi package28. Similar to the result from the PSMC analysis, the wolf population experienced a very mild population growth (1.26-fold increase) that started around 290 000 years ago (Figure 2C). The time of divergence for the wolf and dog populations is inferred to be around 33 000 years ago, where the domestic dog lineage expanded from a population of 4 600 individuals to about 17 500.
In addition to gauging changes in population size, statistical methods can also estimate the rates of exchange of migrants between two populations. The migration rate (2Nm) from the dog lineage to the wolf lineage is estimated to be 0.97, while the other direction (wolves to dogs) is inferred to be 5.02, showing a clear asymmetry in the migration rates29.
Examination of the sequence divergences between the multiple populations using a Markov chain Monte Carlo (MCMC) approach30,31 (Supplementary information, Data S7, Figures S6-S8, Tables S5 and S6) reveals a similar profile for the history between wolves and dogs, which includes a slight growth in the wolf population and an ancient divergence between wolves and dogs (Supplementary information, Data S7 and Table S5). In summary, multiple levels of genetic information (i.e., both joint site frequencies as well as sequence divergence) support an ancient split between dogs and wolves.
The geographical origins of dogs: a single origin in southern East Asia
In order to identify the most probable geographical origin of dogs, we hypothesized that similar to many organisms, the geographical origin of a species holds the greatest genetic diversity, and the global relationship among multiple populations will, in the absence of strong influence of admixture, follow a serial founder model32,33. In the case of dogs, the wild ancestor, the wolf, has been present along the dog throughout Eurasia, implying that intense dog-wolf admixture could possibly have influenced this pattern.
Despite the concern on the confounding effect of wolf/dog gene flow, the TreeMix analysis, F3/F4 test as well as the demographic analysis suggest that gene flow between dogs and wolves is relatively mild. In Supplementary information, Data S8, we review the evidence for dog/wolf gene flow from our study, as well as from multiple previous studies. The combined evidence shows that the migration rates (2Nm) are mostly around one or less (a maximum of five found in the dadi analysis) and that the admixture proportion is normally around 10%, with a maximum of 16% for the Middle East (Supplementary information, Data S8). Low levels of migration are detected between wolves and dogs across Eurasia when the very sensitive D test is used34,35 (Supplementary information, Data S8). Thus, we conclude that while dog-wolf gene flow has occurred throughout history of the domestic dog, it has been at a moderate level and the level of admixture has been relatively similar across Eurasia (Supplementary information, Data S8). Without the strong influence of admixture32, we may assume that genetic diversity is highest at the place of origin and that the global relationship among the multiple populations follows a serial founder model reflecting their dispersal routes33.
It is tempting to draw conclusions about the origin of dogs from the high genetic diversity observed in the Chinese indigenous dogs. However, comparing breed dogs with indigenous dogs at the individual level is likely misleading since most of the differences in genetic diversity are probably caused by recent bottleneck events rather than their distant origin1. Thus, we combine multiple breeds in each region as a group representing the ancestral haplotype pool giving rise to the contemporary dogs of that region. Our analysis shows that dogs from East Asia have the highest genetic diversity (Figure 1E). This suggests that the ancestral population that gave rise to East Asian dogs was much larger than ancestral populations in other regions (e.g., Europe). The linkage disequilibrium pattern also shows the same trend (Figure 1F). Higher levels of genetic diversity in East Asian dogs are also observed in mtDNA and Y chromosome data7,12,36.
Beside group diversity, in the phylogenetic and TreeMix analyses, the deepest node connecting all dogs separates into two clades, one of which is composed of only East Asian dogs, while the other clade includes both East Asian and non-East Asian dogs (Figures 1I and 2A, and Supplementary information, Figure S5). Dogs from Africa and Europe share a most recent common ancestor, which then coalesces with dogs from East Asia (Figures 1I and 2A). Notably, this basal position of East Asia is robust to the levels of migrations between wolves and dogs (Supplementary information, Data S9, Figure S9, and Table S7). The basal position of East Asian dogs is similar to the pattern observed for Africans within human populations37.
In addition to the observations based on group level diversity and the basal phylogenetic position, the PCA pattern also provides supporting evidence for the southern East Asian origin of dogs. As the amount of genetic drift in basal groups is typically lower due to their larger population sizes, we expect them to display a closer genetic relationship with wolves in the PCA plot (Figure 2A). When we simulate a serial founder model that mimics the history of dog domestication, we can easily generate a pattern that is similar to that shown in Figure 1G (see also Supplementary information, Data S10 and Figure S10). Thus, in our analysis, we find dogs with ancestry in southern East Asia to be closest to wolves, and also a geographical distribution of the populations following a clear east-west gradient, indicating serial founder events. It is important to emphasize that admixture between wolves and dogs is unlikely to have created the observed pattern, given that the dog-wolf admixture rate in East Asia is not higher than that seen in other regions (Supplementary information, Data S8).
Having identified southern East Asia as the likely origin of dogs, we asked whether the domestic dog may have originated in more than one region through separate domestication events. In order to test whether multiple origins are compatible with the observed data, we performed simulations mimicking different scenarios (Supplementary information, Data S11 and Figure S11). Our results show that, if there were multiple origins for dogs from separate wolf populations, the descendant populations would tend to reside in separate clusters in the PCA plot, which is in contrast to what we observe (Figure 1G, inset). Thus, that the domestic dog originated multiple times in different geographical areas is not compatible with the observed genetic patterns found in our genome data.
The out of southern East Asia history for the domestic dog
To study the subsequent global history of the dog, we used an MCMC approach to date several important transitional points among the major clades (Figure 2A). Our analysis supports the split between the southern Chinese indigenous dogs and all other dogs across the world around 15 000 years ago, thus indicating a radiation of dogs out of southern East Asia earlier than the origin of agriculture (Supplementary information, Data S7 and node 2 in Figure 2A and 2D)38. After radiating from southern East Asia, possibly following existing human settlements at the time (Supplementary information, Data S12 and Figure S12), the out of southern East Asia lineage spread to the Middle East/Africa and arrived in Europe by about 10 000 years ago (Supplementary information, Data S7; node 3 in Figure 2A and 2D). Notably, one of the out of southern East Asia lineages migrated back to northern China, meeting endemic Asian lineages that had spread from southern East Asia and yielding a series of admixed populations, including the northern Chinese indigenous dogs and the Arctic dog breeds (Figure 2A and 2D).
Several dog breeds from South and Central America (i.e., Chihuahua, the Mexican and Peruvian naked dog) show no signs of admixture, while the Arctic breeds, Alaska Malamute and the Greenland dog, display extensive admixture from the southern Chinese Indigenous lineage39. Possibly, this reflects that the human colonization of the New World occurred in several waves, in which dogs may have followed in different time periods40 (Figure 2D). Using the patterns of the admixture tracks, we estimate that the time of the admixture for the northern Chinese indigenous dogs was quite ancient (around 10 500 years ago, Supplementary information, Data S13 and Figure S13)40. The relatively recent origin of European dogs (i.e., ∼10 000 years) together with this rather ancient admixture suggests that multiple lineages travelled to the Far East from the Middle East/Europe.
Population structure among wolves
Our structure and principal component analyses do not reveal any population substructure among the gray wolves collected for this study (Figure 1D). The high migratory ability of the gray wolf might allow the populations to remain highly homogenous across the eastern part of Eurasia41. A previous study using wolves from the Middle East (Israel), Europe (Croatia) as well as China found genetic differentiation among these wolf populations6. When these three individuals are overlaid on the large PCA plot, the wolves from western Eurasia do not group together with the wolves we collected from eastern Eurasia, and they are genetically closer to dogs (Supplementary information, Data S14 and Figure S14). Given the fact that Middle Eastern wolves generally have more dog admixture6, the observed difference might not represent true population differentiation among wolves. Nevertheless, it is possible that some wolves have recently diverged from each other8, as there is weak isolation between the wolves from eastern and western Eurasia. Explicit testing for potential admixture between wolves and dogs sampled in our study finds evidence of gene flow between wolves and local dog populations in each region, albeit the magnitude is low (Supplementary information, Table S8). Further study on the genetic and geographic relationships between dogs and wolves is one of the important tasks for the community.
Domestication genes
Our analyses indicate that the Chinese indigenous dogs represent an intermediate form between wolves and breed dogs, and they have not experienced intense artificial selection. Analyses of Chinese indigenous dogs therefore allow us to stratify the domestication process in dogs, and investigate the role of positive selection that occurred specifically during the first stage of domestication. Using a statistical method that explicitly models selective sweeps42, we have identified the top 1% of the genome bearing strong statistical evidence of positive selection in the southern Chinese indigenous dogs. In Table 1, we list the categories of genes that show statistical significance by a gene enrichment-based analysis. Groups of genes showing the strongest evidence of positive selection are those related to metabolism and motility, neurological process and perception as well as sexual reproduction (Table 1 and Supplementary information, Data S15, Tables S9 and S10). Genes that seem to have been positively selected in subsequent evolutionary steps, including dog breed formation, are related to the control of developmental processes and to metabolism (see a full discussion of candidate genes involved in transforming wild wolves to dogs in Supplementary information, Data S15).
Among the candidates as positively selected genes in the first stage of dog domestication, a class of genes are related to memory and long-term potentiation (LTP), which is widely considered to be the major cellular mechanism underling learning and memory43. For example, GRIA1 (glutamate receptor, ionotropic, AMPA 1) is an important protein that mediates excitatory synaptic transmission in the central nervous system and plays a key role in hippocampal synaptic LTP and long-term depression (LTD). Interestingly, a suite of other genes, including GRIN2A (glutamate receptor, ionotropic, N-methyl D-aspartate 2A), are also found to be heavily involved in LTP and LTD (Table 1). The large physiological and behavioral changes empowered by these genes may have enabled the transformation of gray wolves to domestic dogs, allowing them to flourish in the human environment.